A Boolean model of the esophagus
![]()
The purpose of a model is to reduce the real thing to something simple enough to be studied free of obscuring detail. This model of the esophagus, as an extreme simplification, reduces it to three elements, the circular, sphincter and longitudinal muscles. The superior constrictor will be ignored. Subsequently I will show that both longitudinal and circular muscle contraction can be either peristaltic or en masse, however this too will be ignored in the model.
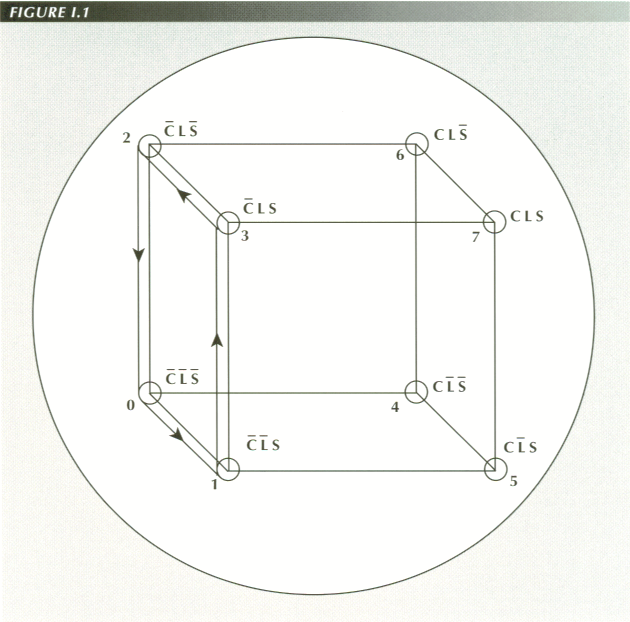
BOOLEAN MODEL OF SWALLOWING LIQUIDS
The state of the esophagus moves along the edges of a cube defined by the 8 possible combinations of relaxation/contraction of its constituent elements: the circular, longitudinal and sphincter muscles
The status of the esophagus at any instant in time can be specified by giving
the state of each muscle. These specifications are achieved by "and-ing" the
three symbols or their negations. Thus, the normal condition of the esophagus
is ~C&~L&S - that is, the longitudinal and circular muscles are relaxed
and the sphincter is contracted.
Detecting the formula for the resting esophagus is easy because it stands still
while we are doing so. We can tell that the sphincter is closed because there
is no reflux, that the LM is relaxed because there is no shortening, hiatal
herniation or tenting of the diaphragm or PEL. The circular muscle state is
less obvious, but with the fluoroscope we can see that there is no peristalsis
going on, a swallow of barium initially meets no resistance when discharged
from the hypopharynx. Manometrically the pressure is zero or negative.
Even at this point, the insight provided by a model enables one to ask some
questions, the answers to which, if they could be found, would not be trivial.
During deglutition the state formula must change from the resting formula. Knowing
the exact sequence in which the state formulas changed during this function
of the esophagus would be interesting. Obviously, there is no reason to exclude
L, the state of the longitudinal muscle, from consideration.
Before attempting to trace the changes in the state formula, a further simplification
will be employed as it will give direction to the search. We can assume they
are there, drop the ampersands and write the resting formula as ~C~LS. In addition,
because its order in the formula already identifies the muscle, we can use the
binary numbers 0 and 1 to stand for the relaxed and contracted states of the
components. Thus simplified, the resting state formula becomes 001.
If these binary numbers are then treated as the x, y and z coordinates of a
point in space,(1) they define the 8 vertices
of a unit cube. This produces a major simplification at once. We can require
that only one of the three muscles changes its state at a time. This is equivalent
to a rule restricting the "state paths" to those from one vertex to an adjacent
vertex. Thus, instead of having 7 possible ways things could change from one
state to another, the possibilities are reduced three. Because the only normal
state node of the organ is 001, a second rule is that all paths on the 3-D cube
must be closed.

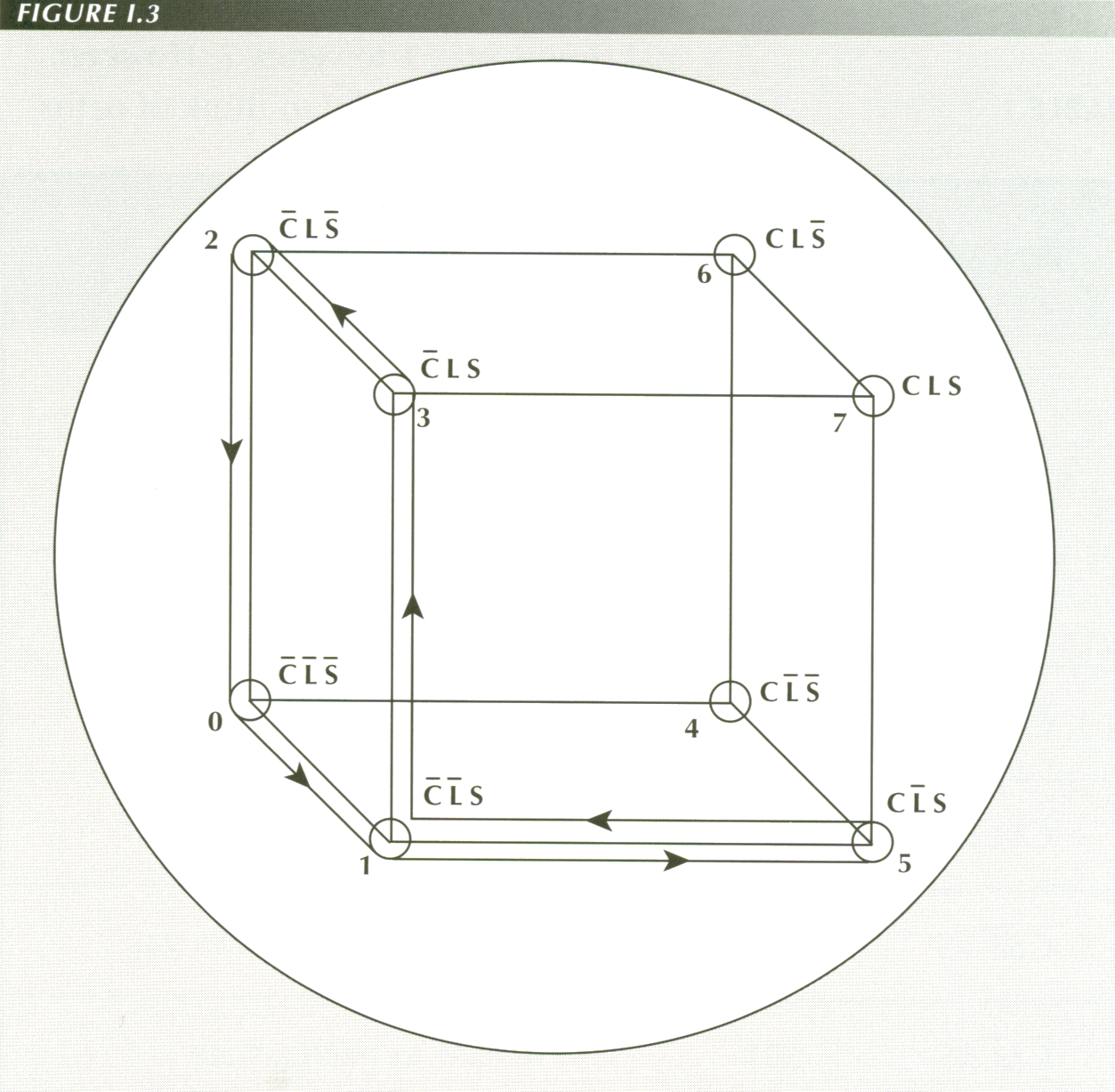
BOOLEAN MODEL OF SWALLOWING AGAINST RESISTANCE
The problem is now reduced to deciding what happens first. That done, one is
ready to look for the next change, one component at a time. Tracing the entire
path in a given subject is not even necessary. It is enough to work out one
transition. The next can be worked out at liesure. A directed observation is
far easier and more likely to be accurate than sizing up the esophageal gestalt.
In addition, the longitudinal muscle (L) now becomes an essential part of the
picture at least as important as the other components. Simply because it does
not affect a manometer or a balloon is no reason for ignoring it.
![]()
Another consequence of the model is to emphasize that the esophagus is not a single-purpose organ designed only for swallowing. This is because there are a great many paths from vertex to vertex that can be traced on a 3-dimensional cube. Here is a clue to the ability of the organ to swallow liquids via a different path than it uses for solids or when swallowing against resistance or in the upside down position. We can now see that esophageal speech might have a still different path. Deploying its resources in varied sequences is the means by which the esophagus carries out its multiple functions - swallowing liquids and solids, belching, gagging, vomiting.
In principal, it would be also be anticipated that various malfunctions could
cause interruptions of the smooth transitions along normal pathways or that
arrests could occur - possibly at different vertex of the cube. When these very
specific questions have been answered, we should have a much more detailed knowledge
of the diverse functions of the organ.
Fluoroscopic observation is generally sufficient to map the paths of changing
state patterns with the aid of this search algorithm. In principle, however,
it should be possible to develop instrumentation to detect all three types of
muscle contraction simultaneously. A computer could constantly monitor the changing
patterns with a high degree of accuracy.
I will trace the state pattern in several modes of esophageal activity. By
treating the 3-bit binary coordinates of the vertices as octal numbers and using
them as labels for the vertices they can be referred to more conveniently. Thus,
111 becomes 7, 001 translates to 1, etc.
BOOLEAN MODEL OF BELCHING
The contraction of L changes the formula to ~CLS or 011 = 3 and the subsequent
sphincter relaxation to ~L (010) = 2. A rapid alternation among nodes 1, 2 and
3 then occurs with each subsequent swallow. The graph never gets off the not-C
plane.
Swallowing against resistance, whether it be with solid food, in the upside
down position or during a Valsalva effort causes the path to leave the left
(not-C) plane and involve the circular muscle. The first event is active peristalsis
(001 to 101). Peristalsis gradually "latches" the longitudinal muscle (101 to
111) causing the sphincter to yield (110). Just before the peristaltic wave
reaches the sphincter, the longitudinal muscle relaxes (110 to 100), the ring
of circular muscle contraction becomes the sphincter (100 to 101) and
so vanishes (101 to 001). The path traced is 1, 5, 7, 6, 4, 5, 1.
As there are three possible transitions from a given vertex, and swallowing
involves 6 transitions, there are 33 or 729 possible sequences of
which only one is appropriate to swallowing against resistance.
Comparison of the graphs of the two swallowing modes shows strikingly different
pathways. Note that the state formula changes one bit at a time, and that it
is difficult to leave the designated path without the swallow aborting. Both
activities are highly mechanical acts that the model represents quite faithfully.
One might say that LMC is essentially concurrent with the peristaltic wave and
ought to be represented as a path on the diagonal from vertex 1 to vertex 7.
However, it seems more helpful to think of paths confined to the edges of the
cube between vertices even if close temporal relationships or overlapping activity
warp the picture.
For belching the following schema is easily detected:
| Formula | Node | Esophageal state |
| ~C&~L&S | 001 | Resting state |
| ~C&L&S | 011 | Longitudinal muscle contracts |
| ~C&L&~S | 010 | Sphincter opens, gas escapes |
| ~C&~L&~S | 000 | Longitudinal muscle relaxes |
| ~C&~L&S | 001 | Sphincter closes |
| C&&S | 101 | Circular muscle evacuates gas |
| &&S | 001 | CM relaxes to resting state |
![]()
It is evident from inspection of the various state-path diagrams that they
are not restricted to a single plane but move in all 3 "dimensions." This highlights
a remarkable aspect of esophageal physiology to date: it has been largely restricted
to the CS plane. That is, it is fundamentally 2-dimensional.
If this handicap is accepted, we are as limited in our comprehension as were
the Flatlanders of Edward Abbott,(2) who could
be imprisoned by circumscribing them with a line because they could not conceive
of a third dimension. The available balloons, manometers and transducers are
Flatlander instrumentsas they see only the circular and sphincter muscles.
This has serious consequences even for careful laboratory research. No matter
how meticulously LES pressure and peristaltic wave pressure are measured or
sequenced under various experimental conditions, the interactions between stimulus
and response may be happening in a plane that is invisible to the experimenter
who is not also tracking LM function. Unfortunately, instrumentation has not
been developed to measure LM contraction conveniently in the intact subject.
Another aspect of esophageal physiology that can be read from the model is
the fact that, with one exception, the organ normally never remains indefinitely
at a node. It occupies them only in transit. The exception, of course, is the
resting state - (001). Permanent occupation of another vertex is pathologic.
Myotonia dystrophica, for example, seems to arrest the organ at 010 as does
scleroderma.
The various formulas that occur can be listed and "or-ed" together and so manipulated
with operations of the sentential calculus of mathematical logic(3)
to yield a result that I will subsequently show to be supported by radiological
and clinical observations. It might be called the Fundamental Law of the Esophagus:
~S <-->L
That is, the sphincter is open if and only if the LM is contracted.
SUMMARY
Because it ignores the function of the longitudinal muscle, the prevailing esophageal paradigm is 2-dimensional. A 3-D model reveals several rules of esophageal physiology, the most important of which is the rule that the longitudinal muscle and the sphincter are never contracted simultaneously. This is equivalent to the assertion that the LM opens the sphincter. The vastly increased number of possible state sequences together with alternative peristaltic and en masse modes of contraction account for the ability of the esophagus to carry out a variety of functions.
![]()
References
Last Updated July 25 2007 by David PJ Stiennon
1. . Flegg, H.G., Boolean Algebra and its Applications, John Wiley & Sons, New York, 1964.
2. . Abbott, Edward, The Flatlanders, Dover, New York, 1952.
3. . Hilbert, D. and Ackerman, W., Principles of Mathematical Logic, Ed. Luce, Robert E., Chelsea Publishing Company, New York, 1950.